




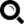
Magyarok Világszövetsége — Magyarságtudományi Füzetek
Sejti örökléstani fogalmak
A földi élet alapelemei a sejtek. A sejtek alapvetően két nagy részből állnak: a sejti élet mechanizmusát biztosító plazma és tartozékai, valamint az életjelenségeket meghatározó, a sejt valamennyi jellegét meghatározó örökletes anyagból. Az állati szervezetek örökletes anyagát, a sejti átörökítő anyagot (idegen szóval géneket) két alakban találjuk meg: az egyik a sejti mechanizmus része, amit a mitokondrium tartalmaz – és ez biztosítja a sejtlégzés mechanizmusát, – míg a másik, a sejt egészére vonatkozó minden örökletes anyag a sejt magjában helyezkedik el. Itt pedig a szervezet minőségétől függő számban találunk bázikus színezékkel könnyen megfesthető, azaz savas jellegű részeket, idegen szóval: kromoszómákat, melyek az átöröklés anyagát dezoxi-ribo-nukleinsavak (továbbiakban DNS-sel jelöljük) alakjában tartalmazzák.
A kétivarú szaporodásnál a sejtmag örökletes állományát fele részben a nőstény, fele részben a hím biztosítja. Ezért a kromoszómák párban vannak, melynek egyik eleme a hímtől, a másik a nősténytől származik és a sejtfejlődés során, a szervezet kiépülésekor a sejt ezek közül véletlenszerűen válogatva az egyiket használja fel. Van azonban egy kromoszómapár, ami valójában nem pár, csakis a nősténynél van valódi párban, a hímnél annak egyik eleme a másiktól különbözik. Embereknél ezt a kromoszóma párt X és Y kromoszómának nevezik. X a nősténynél valóban párban van, mert két szülőtől származó X elemet tartalmaz, a hímnél azonban a nemiséget az egyik elempár, az általában kisebb, az Y jelű összetevője határozza meg.
A kromoszómákon helyezkednek el a tulajdonságot meghatározó elemek – idegen szóval: gének – és ezekből az embernél összességében mintegy 30 ezer van. A sejti átörökítők ezen elemeinek átörökítést biztosító részei DNS-ből állnak, melyeket némi fehérje és zsíralkotó köt össze génné és tovább kromoszómává.
Kétivarú szaporodásnál a hímek az Y sejti árörökítőjüket kizárólag a felmenő hímtől kapják, ezért az Y sejti átörökítők elemzése betekintést nyújthat a hím ágon való örökösödési kapcsolatláncba. Különösen igaz ez azért, mert az Y sejti átörökítő egyik szakasza már nem hordoz az életre vonatkozó információt, ezért az utód életének veszélyeztetése nélkül megváltozhat – és az idők során meg is változik. A változások az utódokra átadódnak, ezért a változások elemzésével nyomon követhető az apa-fiú kapcsolatok lánca. Ez szolgálja a férfiági családfák felderítését – és a modern kori genetika képessé vált a változások kimutatására.
A sejti mechanizmust minden esetben a nőstény biztosítja, így a sejtlégzés mechanizmusa is tőle ered. A sejtlégzésnek is van sejti átörökítője és ezt a sejtmagtól megkülönböztetendően – idegen szóval – mitokondrium sejti átörökítőnek, a lényegét alkotó vegyi anyagot pedig mitokondrium-DNS-nek nevezzük és a továbbiakban mtDNS-sel jelöljük. Ennek változásait vizsgálva a női felmenő ’családfa’ tanulmányozható.
DNS vizsgálatok
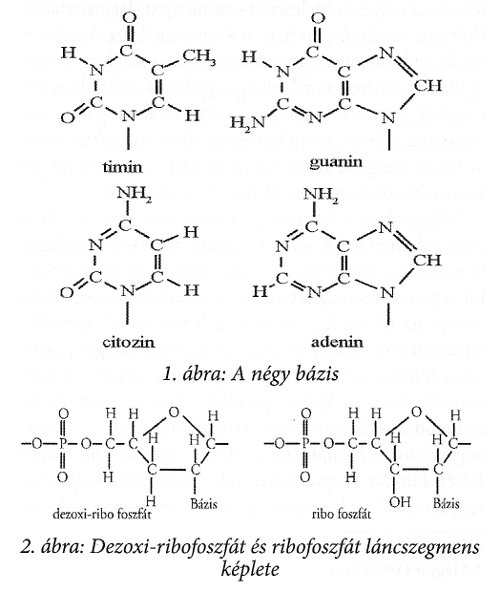
A dezoxi-ribonukleinsavak három alkotórészből állnak: két foszforsavból és a dezoxiribózból álló makromolekuláris lánc kettős hélixet alkot, melyeket a ribózra kapcsolódó bázisok tartanak egybe hidrogén hidakkal. Egyetlen molekulaláncú hélixet képez a poli-ribofoszfát lánc a hozzá kapcsolt bázisokkal, melyben a DNS dezoxiribóz molekulája helyett annak oxidált alakja, a ribóz vesz részt. Ezt a láncot RNS-sel jelöljük. A ribfoszfát és a dezoxiribofoszfát szegmens molekulaszerkezetét a 2. ábra szemlélteti.
A DNS-ek bázisa alapvetően négy fajta lehet (1. ábra), melyek szigorúan párban alkotják a DNS-t: a guanin a citozinnel, az adenin pedig a timinnel van párban. A mtDNS-nél az egyik bázist annak metil csoportot tartalmazó változata helyettesíti (citozint az uracil). A sejti átörökítést a 3. ábra szemléleti.
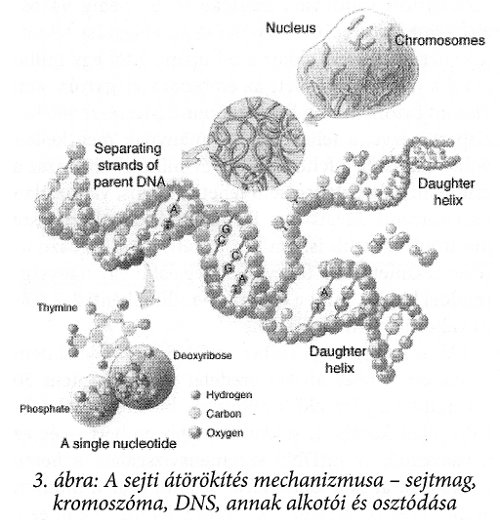
A sejti tulajdonságot – az egyed teljes szervezeti fölépítését, működési mechanizmusát stb. – az egyes DNS molekulákon elhelyezkedő bázisok sorrendje határozza meg. Minthogy a DNS kettőshelix alakban létezik a szervezetben, a bázisai kívülről nem láthatók, szigorúan védettek. A sejti szaporodás mechanizmusa biztosítja, hogy szaporodáskor a DNS kettőshelix kétfelé bomlik és ezen közben a láncnak megfelelő tükörlánc épül mellé, azaz két teljesen azonos DNS lánc keletkezik.
A tulajdonságok kiolvasásakor ugyanez játszódik le, de akkor a második lánc nem dezoxi-ribózt, hanem annak oxidált változatát, ribózt tartalmaz. Ez az apró különbség elegendő ahhoz, hogy az így keletkezett poli-ribofoszfát lánc bázisai (RNS) az információt hordozó bázisait a helix külső felületén tartalmazza, és ezért biokatalizátorként viselkedhet. De erről készülnek a sejtet alkotó, a sejtben lejátszódó reakciókat katalizáló fehérje molekulák is. Három egymást követő bázis képvisel egy aminosavat az RNS-ről készített fehérjéknél.
E rövid, sejti átörökítő tulajdonságot bemutató összefoglalónk lényegi mondandója az, hogy a sejti átörökítést biztosító molekulák semmi mást nem kódolnak, mint a sejtet fölépítő fehérjéket, illetve azokban lejátszódó biokémiai reakciók katalizátorait. Hibás minden olyan felfogás, hogy a sejti átörökítők módosításával az ember oly mértékben beavatkozhat az örökletes folyamatokba, amint azt számos eugenetikus elképzeli.
A másik következtetésünk az, hogy a sejt jövőjét nem befolyásoló szakaszokon bekövetkezett bázissorrend változását ma már elemezhetjük, és ez alapján következtethetünk emberek, embercsoportok eredetére, kapcsolatára – visszamenőleg a legtávolabbi múltba, egészen az emberiség kialakulásáig, sőt, azon is túl. Ezt két ágon, a csak hímről hímre átadott Y kromoszóma egy masszív nem védett szakaszán, illetve az mtDNS egy viszonylag rövid (440-2000 között változó) szakaszán vizsgálhatjuk.10
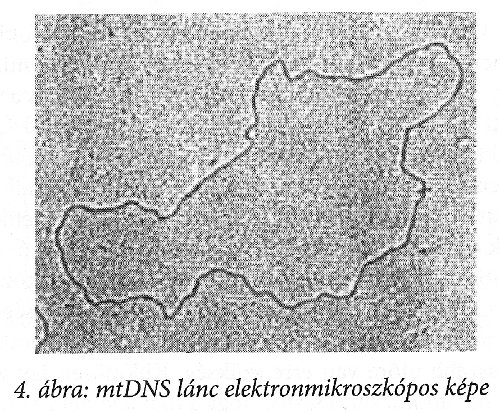
A 4. ábra az mtDNS elektronmikroszkópos képe. A DNS végtelenített szálat képez, melynek vastagsága 1 nm, ahol az egyenes szakasz nagysága megfelel a változásra alkalmas szakasz becsült nagyságának. Ez a DNS 16569 bázispárt tartalmaz, s ebből kb. 1200 változhat meg a mitokondrium működésének veszélye nélkül. Valójában 440 bázispárból álló szegmenst vizsgálnak.
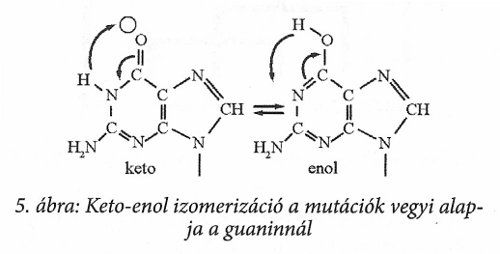
A változást mutációnak nevezzük, amikor is az 5. ábrán bemutatott módon a guanin bázis jellege megváltozik és hidrogén befogadóból hidrogén adóvá válik, amitől a vele szemben a szaporodáskor beépülő bázis a citozin helyett megfelelő szerkezetű bázissal, timinnel helyettesítődik. Ha a timin változott, akkor adenin helyett guanin lesz a párja. Minden további szaporodásnál a megfelelő pár épül be, így az eredeti adenin helyett majd guanint és citozin helyett timint találunk azon a helyen. A guanin és a timin alkalmas a változásra, a másik két bázis szerkezete állandó.
A mutáció azonban meglehetősen ritka. Genetikusok azt állítják, hogy egy-egy mutáció létrejöttéhez 10 évezred is szükséges, más megfogalmazás szerint évmillióként a bázisoknak 2-2,4%-a változhat meg.11 Ebből ún. genetikai órát állítottak fel és ennek alapján számítják, hogy egy-egy mutáció mikor következhetett be.
Mitokondrium DNS vizsgálatok
Az mtDNS 440 szegmensét vizsgálják a öröklési láncnál. Évmillióként a teljes mitokondriumban mintegy 360 bázis változhat meg. Ámde ha a változás a védett szegmensen történik, akkor az a mitokondrium kiiktatódik a használatból – sejtenként ezernél több is jelen lehet! – a 440-es szegmensen pedig ez évmillióként 4-5 változást eredményezhet. Mégis, a genetikusok ennél sokkal gyorsabb változást tételeznek, és azt állítják, hogy 10 évezred szükséges egy-egy változáshoz.12 Holott ha valóban statisztikus valószínűséggel történik a változás, akkor ennél egy nagyságrenddel hosszabb időre van erre szükség. Különösen így van ez, ha tudomásul vesszük, hogy a mutáció nem az idővel, hanem a szaporodási esetekkel arányos és ezért alacsony népesség mellett sokkal lassabban történik egy–egy mutáció, mint igen nagyszámú esetben.
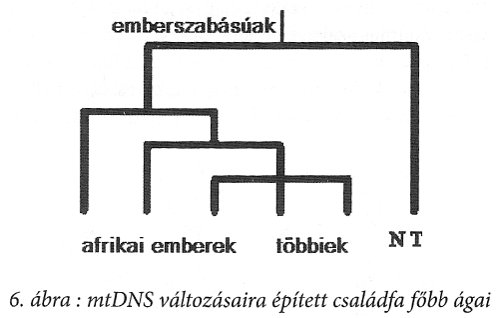
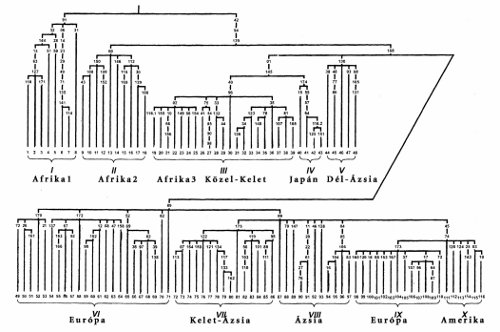
Az emberiség több tízezer tagjánál vizsgálták meg a mitokondrium ezen szakaszát. Megállapították, hogy a teljes emberiségre nézve a változás legfeljebb 35 bázist érint.13 Az átlagos különbség ember és ember között 11 bázist jelent. A változásokra fölépített ‘családfát’ a 6. ábra szemlélteti.14 Látható, hogy a változások zöme az afrikai emberhez köthető, míg az összes többi ennek csupán kései kiágazását jelenti – és ezzel az emberiségnek az afrikai származása szinte bizonyított is. A gond az időrenddel van. Az első elágazást ugyanis 150 évezreddel a jelen előttre, a nem afrikaiakét pedig mintegy 50 évezreddel ezelőttre teszik a genetikusok.15
Az időrenddel azonban több gond is van. Mindenekelőtt, azért is, mert ismeretes a csimpánzokhoz viszonyított változások mértéke is. Ez pedig 94 bázist érint.16 Ha a 10 évezredenkénti elágazást tekintjük mértékadónak, akkor a csimpánzoktól egy millió évvel a jelen előtt kellett az embernek elágaznia, ami viszont bizonyítottan hibás időpont. Másrészt 440 bázispárt nézve, a teljes bázisállománynak meg kellett volna változnia a feltételezett 5 évmillión belül, azaz a csimpánzokkal elvileg sem lehetne közös, változatlan bázissorrend. Mégis van. De hasonló következtetésre jutottunk föntebb is, amikor a statisztikai valószínűséget tekintettük: a tíz évezred legalább egy nagyságrenddel kisebb, mint a bázisok évmilliónkénti 2-2,4%-os változási várható értéke.
Ekkor pedig a mai ember mtDNS szekvenciái nem a modern ember afrikai eredetét igazolják, nem 50 évezreddel a jelen előtti kijövetelt, hanem annál sokkal-sokkal korábbit, sokkal idősebb embert – és ez a valószínű. A mtDNS szegmensvizsgálata a homo erectusnak afrikai kiáramlását jelenti, azaz a modern ember ezek összességének a tükre. Kínos? Nem: ez a tudomány!
A 6. ábrán a modern embertől elkülönülten ábrázolják a neandervölgyi ember mtDNS szekvenciáját. Innen azt igyekeznek kiolvasni, hogy a neandervölgyi ember kihalt, nem lehet a modern ember elődje. Néhány neandervölgyi ember csontjából sikerült kinyerni DNS szegmenseket – ami önmagában véve is izgalmas, hiszen a szabad DNS néhány évszázad alatt általában elbomlik. Emberi csontokból kinyert kollagén alkalmas a vércsoport meghatározására, de a DNS vizsgálatok eredménye rendkívül bizonytalan. Mégis azt állapították meg, hogy az átlagos ember mtDNS-étől 35 szegmensben különbözik a neandervölgyi emberé17 – és ez nem nagyobb távolság, mint az emberiség teljes szegmenskülönbségének a nagysága. Mindössze 3 neandervölgyi ember adatai ‘ismertek’ – ha elfogadjuk a rizikós eredményt.
A mtDNS szegmens változásait családfába rögzítve állította Sykes (2001),18 hogy az összes mai ember anyai őse egyetlen asszony volt és annak 7 ágra szakadt utódcsaládja ismerhető fel. Ez az ősanya, szerinte, 150 évezrede, Afrikában élt.
Ám ha az időrendre tekintünk, akkor bizony egy tízes szorzót fel kell tételeznünk, azaz ez az ősanya nem 150 évezrede, hanem mintegy másfél évmillióval ezelőtt élhetett Afrikában. Ez az időpont a régészeti adatok alapján értelmezhető is, a 150 évezredesnél komoly gondok vannak (lásd később).
Y kromoszóma vizsgálatok
Az Y kromoszómának nemcsak ilyen rövid szakasza, mint az mtDNS-nél, hanem sokkal több bázist tartalmazó szakasza változhat szabadon, ezért ennek elemzése sokkal finomabb tagoltságot enged meg. A jellegzetes változást kísérő egyéb változásokkal a mutációk számának sokasága lehet még jelen, ezért a változások szélességéből itt is időrendet lehet megállapítani. A lényeget tekintve elfogadható, hogy a szélesebb eloszlás régebbi mutációt jelent, míg a szűkebb közelebbit – de az abszolút időskála itt is kétséges, jóllehet a vonatkozó irodalom azt bőségesen ontja.
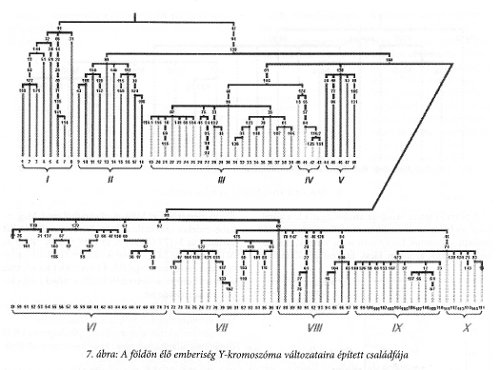
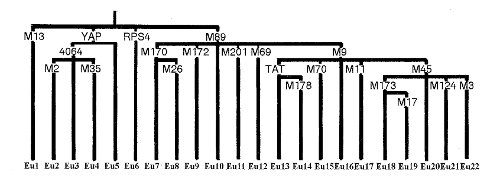
A 7. ábrán az Y-kromoszóma szabadon változó szakaszáról megállapított gén-családfa látható Underhil és munkatársai alapján.19 A családfa szerkezet főbb vonalaiban megegyezik a mtDNS családfájával, csak annál jóval bővebb, részletesebb. Az egyes csoportok helyi eloszlást jelentenek. Az I és II kizárólag Afrikában létezik. Ebből ágazik le az M168 jelű ág, melynek egy része ugyancsak Afrikát jelenti (III bal fele), a III-as jobb felét a Közel-Keleten, míg a IV jelzésűt Japánban, és az V-t Dél-Ázsiában találhatjuk meg. Velük párhuzamos a több részre szakadó VI-X szakasz. A VI és a IX jelzésűek az európai előfordulások, VII Kelet-Ázsia, VIII Ázsia, X pedig Amerika jellemzője. Ezek a csoportok a mai emberre vonatkoznak, ami azt jelenti, hogy a megfelelő osztályozás a zömre, de nem minden egyedi embere vonatkozik, hiszen az emberiség keveredettsége manapság óriási.
Az Y-kromoszóma családfája úgy szintén egyetlen őshöz vezet, de ez az ősapa nem akkor és nem ott élt, ahol a mtDNS-ből kikövetkeztetett ősanya.
Számunkra itt két variánscsoport nagyon érdekes. Az egyik az M89-es VI-os csoportja, melyhez az európai emberek sejti átörökítő variánsa tartozik, a másik csoportban találjuk meg Ázsia és Amerika variánsait és egyben Európa leggyakoribb csoportját is, a IX jelzésűt. A főág az M89-es, az al-ágak fő alkotója az M9, amiből az ázsiai, az európai és az amerikai ember variánsa ágazik ki. Külön érdekes, hogy az M9 külön alágát képezi az M45, és ebből ágazik ki az európai és az amerikai variáns (X).
Az alapközlemény Európát érintette és az európai változatok eloszlását ábrázoljuk a 8. ábrán.20

A génfa fő ága az M168-as, amiből ágazik ki a közel-keleti és bizonyos mértékben az afrikai ág is. Az M89 melletti változatok Európában jelen vannak, százalékos mértékük azonban kicsi, Európa génállománya döntő mértékben az M89-es változatból ered. Még inkább érdekes számunkra, hogy Európa uralkodó két variánsa az M173 (Eu18) és az M17 (Eu19). Mind az M173, mind az M17 a genetikusok eredeti kormeghatározása esetében is legalább 40 évezredet jelent, azaz nem az Európába beáramló későbbi népek, népcsoportok hozták ide be.
Az európai variánsok egyes népek közötti megosztását számszerűen is közöljük a 1. táblázatban.
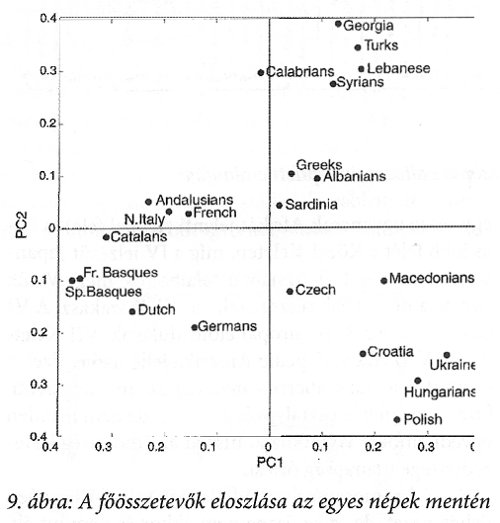
A táblázat adatait matematikailag feldolgozva két fő összetevőt állapítottak meg. Ezek koefficienseit ábrázolva a 9. ábrát kapták a kutatók.

A két főösszetevő eloszlása alapján az egyes népek négy nagy csoportba sorolhatók. Az ábra jobb felső részén Kaukázus környékiek helyezkednek el. Ennek a mezőnek az alsó részén a Földközi-tengeriek, közvetlenül mellettük a baloldali mező alján a közeli nyugat-európaiak találhatók szoros kapcsolatban a mező alsó szegmensén elhelyezkedő többi nyugat-európaival. A negyedik meglehetősen szoros csoport a jobb mező alján található. Ezek középpontjában vagyunk mi magyarok és lengyelek, ukránok és horvátok vesznek körül minket. A csehek és macedónok a nyugat-dél-európaiak felé találhatók.
Nagyon fontos még a fő összetevők súlyának a Közel-Kelettől való távolságában ábrázolt függése, amit a 10. ábra szemléltet.
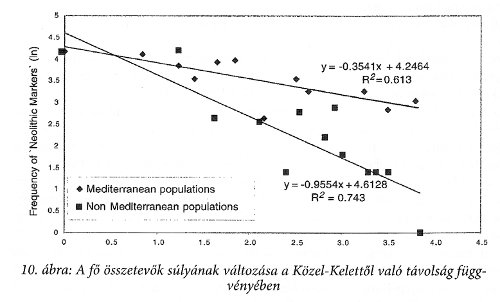
Más a távolságfüggés a mediterrán körzet népeire és megint más Európa belsejének népeire vonatkozóan. Ez az ábra egyértelműen igazolja, hogy az ún. mezőgazdaság-elterjedést nem kísérte a közel-keleti sejtátörökítők jelzőjének az elterjedése, azaz Európa valójában nem a mezőgazdaság terjedésével népesedett be. Ez a mai magyarság eredetére nézve kulcsfontosságú adat!
A 11. ábra a génlánc főelemeinek eurázsiai eloszlását szemlélteti.21 A meghatározás szerint az 1-es a YAP, ami nem tartalmazza az M130-at és így tovább, a 2-es az M130, a 3-as az M89, a 4-es az M9, az 5-ös az M45, a 6-os az M173 és a 7-es az M17.
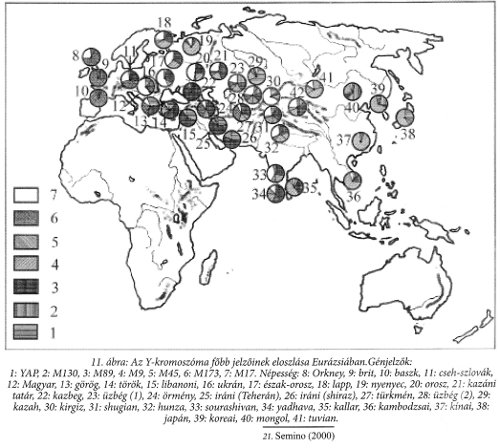
A magyarokra 60% M17, 13,3% M173 a jellemző, hozzájuk legközelebb a lengyelek állnak (56,4 % M17), majd az ukránok (54 % M17), s Magyarországtól távolodva az M17 részaránya rohamosan csökken. Nyugati irányban az M173 részarány nő meg, déli irányban pedig a nem-európai variánsoké.
További érdekes jelenségekre figyelhetünk fel, ha figyelmünket immár Ázsiára szegezzük. Mindenképp úgy tűnik, hogy az M89 a Kaukázus környékét jellemző ősvariáns, míg az M130 halad délnek és fedi le Dél-Ázsiát.
Figyelemreméltó ugyanakkor, hogy az M9 erősen megnő, majd uralkodóvá válik kelet felé, mely alól a japánok képeznek kivételt, akiknél az M89-et megelőző, illetve annak szomszéd ága szolgáltatja a jellemző hányadot.
A sejti átörökítők változatainak térbeli eloszlása az embernek Eurázsiában való elterjedése útját és viszonylagos idejét jelöli ki. Nagyon fontos innen, hogy a YAP a közel-keleti gabonatermeléssel párhuzamosan került be Európába és eloszlása nem terjed ki Európa déli részén felül, nem terjed ki sem Közép-Európára, sem északabbra, nyugatabbra. A két fő európai változat, az M173 (Eu18) és az M17 (Eu19) pedig keleti-nyugati megosztást jelez. Számunkra az a fontos, hogy a magyar lakosságban, a Kárpát-medence lakosságában ez az uralkodó változat 60%-ban van jelen és hozzánk legközelebb az ukránok, a lengyelek, majd a horvátok állnak. Ugyanakkor a magyarországi lakosságban 13,3%-ban van jelen az M173 és a maradékban 8,9%-ban a közel-keleti gabonatermelők változata.
Az Távol-Keletet jellemző M9 európaitól független változata a TAT, mely az észak-kelet-európai népek (finnugorok) jellemző változata, viszont a magyar lakosságból teljességgel hiányzik.
Ide kívánkoznak a legújabb magyarországi mtDNS vizsgálatok.22 Ezek nemcsak a mai lakosok, hanem a honfoglalás kori temetők csontállományára is kiterjedtek és arra a megdöbbentő eredményre vezettek, hogy a honfoglaláskor a Kárpát-medence korábbi, európai eredetű lakói alkották a lakosság zömét, az ún. ‘honfoglalók’ elenyésző kisebbséget jelentettek. Ennek a fentiekkel egyetemben a továbbiakban komoly jelentősége lesz.
Az a tény, hogy mind a férfi, mind a női ágon egy személyhez jut el az örökléstan, nem jelenti azt, hogy egyetlen férfi, vagy egyetlen nő az emberiség egyetlen őse. Ha egy férfinek nincsenek fiú utódai, az ő férfi vérvonala ott befejeződik. Ha egy nőnek nincsenek leány utódai, ez ugyanezt jelenti a női ágon. De a férfinek lehetnek leány, a nőnek férfi utódai, azaz az örökletes tulajdonságok, ha nem is nem-specifikusan, de tovább élnek. Visszafele csak férfi, vagy csak női ágon eljuthatunk valakihez, akinél feljebb nincs, de minden embernek van apja és anyja és ez nem egyenes ágat, hanem hálózatot jelent.
Testi tulajdonságokat tekintve meg rendkívül gyakori, hogy a házaspár fia az anyja, lánya pedig az apja testi jellemzőit örökli.
Vércsoport vizsgálatok
Az emberi vércsoport örökletesen meghatározott. Vagy az apa, vagy az anya vércsoportját örökli a gyermek, harmadik lehetőség nincs.
Az emberiség vércsoportja azonban nem egységes. Ebben is mutációkat találunk, melynek eredményeként 4 fő vércsoport alakult ki az emberiségnél. Az emberiség eredeti vércsoportja a nullás lehetett. Ennek uralkodó mértékével találkozunk Afrikában, Ausztráliában és az amerikai kontinensen. Majd ebből mutációval keletkezett az A és a B, melyeket a legnagyobb sűrűségben a Kaukázus (A) és Mongólia (B) környezetében találjuk meg, azaz vélhetően az ottani emberben történt a mutáció. Az A és B vércsoporttal szemben a nullás vércsoportúak már ellenszérummal rendelkeznek, ami azt jelentheti, hogy a mutáció nagyon régi. A negyedik vércsoport, az AB, és föltehetően legutóbbi időkben keletkezett. Korábban az AB-t az A és a B kombinációjának vélték, mert valamennyi másik vércsoportú embertől kaphat vért, azaz nincs vele szemben ellenszéruma. Viszont a nullás vércsoporthoz való viszonya és az A valamint a B vércsoportnak a nullával való összeférhetetlensége jelzi, hogy ez a mutáció messze nem olyan régi, mint az A és B. De hogy mikoriak és hol jöttek létre? Csupán feltételezésekkel élhetünk.
A vércsoportok ismerete az I. világháború óta nagyon fontossá vált, ezért hihetetlen széles körbeli adatok ismertek. Dr. Nagy Ákos szerológus feldolgozta ezeket az adatokat és megállapította, hogy egyes népcsoportokon és területeken belül a vércsoportok egymáshoz mért aránya meglehetősen állandó. Európában 45%-os aránnyal a leggyakoribb az A vércsoport, ezt követi 35%-os aránnyal a nullás vércsoport, majd 20% körüli a B vércsoport és elenyésző, <5% körüli az AB vércsoport résaránya.
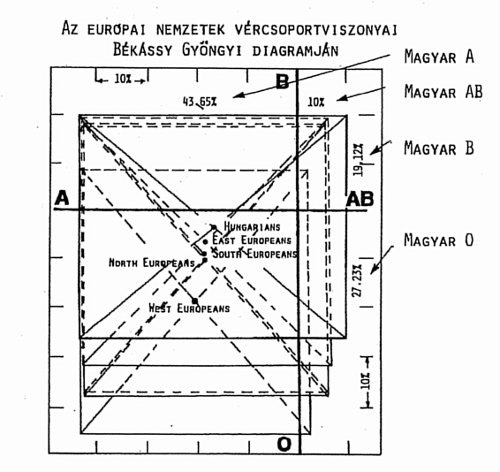
A magyarországi és a világ Magyarországról származó népességénél azonban a nullás vércsoport rovására sokkal magasabb az AB vércsoport aránya (25%). A vércsoportok arányát – Békássy Gabriella ötlete alapján – egy A-AB/B-0 négyszögdiagramban ábrázolva,23 a képezett négyszögek középpontjának egymáshoz viszonyított távolsága jellegzetes népesség jellemző (12. ábra).
Az angol nyelvű ábrán a magyarok (Hungarians) helyzetét hasonlította össze a szerző a kelet- (East Europeans), a dél- (South Europeans), az észak- (North Europeans), és a nyugat-európaiakkal (West Europeans). Az ábrán jól látszik, hogy amíg a kelet-európaiak vércsoport jellemzője a legközelebbi a magyarhoz, a nyugatiaké a legtávolabbi. Ez megfelel a sejti átörökítők elemzésből levont következtetéseknek. A magyarországi vércsoport arányokat külön számszerűen is feltünteti a diagram.

Számos korai temető csontanyagának vércsoportját is megvizsgálták. Az a megnyugtató eredmény jött ki, hogy amint a genetikai variánsok, úgy a vércsoportok aránya is a területen meglehetősen állandó. A területre bejött újabb lakosság aránya az őslakosokétól magabiztosan megkülönböztethető. Ebből az is kiderült, hogy Európába a B vércsoport nem a népvándorlások korában érkezett, hanem korábban is már komoly mértékben jelen volt. Egyébként a B vércsoport Európában keletről nyugatra haladva csökken. Jellemző számnak tekinthető a B és AB vércsoportok részarányainak az összege.
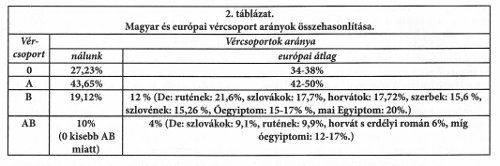
A temetők vércsoport elemzése a sejti átörökítő variánsok eloszlásával összhangban a magyarság őseurópai jellegét igazolja.
Embertani vizsgálatok
Az örökletes jellemzők vizsgálata után vegyük elő az öröklés külső jegyeit: mit is mond az embertan? Itt különösen a csontmaradványok jelentősek számunkra, hiszen az eltemetett holtak csontja kedvező körülmények között százezer évekig vizsgálható állapotban, azaz ’épen’ megmarad, így a történelem előtti ember testi fölépítése, koponya alakja, méretarányai, csontjainak vastagsága kényelmesen tanulmányozható, osztályozható.
Az embertípusok alcsoportra osztásában a testmagasság, a test robusztussága és a koponya jellemzői számítanak meghatározónak. A jelenleg élő ember tipizálásához fölhasználják még a bőr, a haj és a szem színét, ill. a szőrzetet, annak mennyiségét és göndörítettségét.24 Ez utóbbiak a temetkezési helyekről előkerült csontvázak osztályozásában általában nem játszanak szerepet, jóllehet néhány mumifikálódott tetemnél ezek is megállapíthatók. Ugyancsak a vércsoport és az egyre nagyobb mértékben megismerhető sejti örökléstani sajátságok is segítenek ma már a csoportosításban, de a történelem előtti embernél ezek még szintén nem nagyon jöhetnek számításba, jóllehet csonttöredékek alapján újabban már ezek a jellemzők is meghatározhatók.25
Az emberek osztályozásánál a testarányok és a koponyán mérhető arányok a legjelentősebbek. A testarányokban komoly szerep jut annak az éghajlatnak, melyen az ember él, tevékenykedik. Alapvetően hideg- és meleg övi ember típusokat különböztethetünk meg.
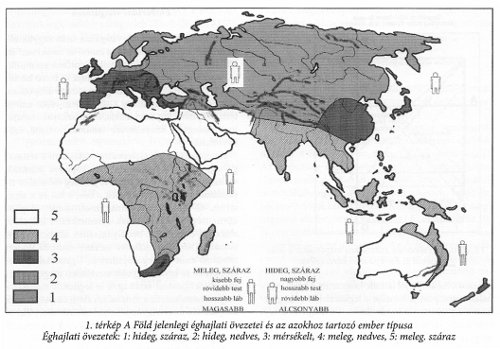
A korai ember egészen a neandervölgyi és a modern ember koráig kerülte a száraz, kifejezetten hideg éghajlatú helyeket, jóllehet, már a legkorábbi eurázsiai emberi telepek is inkább a hideg, semmint a mérsékelt, vagy meleg égövi területről ismertek (Swanscomb, Vértesszőlős, Zhoukoudian). Az ember testalkata igazodott az éghajlati körülményekhez. A meleg övi ember feje általában kisebb, mint a hideg övié, teste is kisebb, lába viszont hosszabb, teste összességében magasabb. Az antropológusok ezt arra vezetik vissza, hogy a meleg övben kisebb hőelnyelő-képességű testet a talajtól távolabbra kell vinni, mint hideg övben, ahol viszont a fejet kell védeni a nagyobb lehűléstől. A gond azonban az, hogy mindez mennyi idő alatt tud kialakulni? Egy meleg övi ember, ha hideg övezetbe kerül, vagy egy hideg övi a melegbe, mennyi idő alatt alkalmazkodik ehhez? Ausztrália első lakói hideg övi emberek voltak és létük közel 30 évezrede alatt testalkatuk nem sokat változott. Az 1. térkép nem csak az éghajlati övezeteket, hanem az azokhoz rendelhető ember testalkatát is bemutatja.
A tapasztalat az, hogy az Eurázsiából ismert legkorábbi modern ember hideg övi testalkattal rendelkezett. De hideg övi testalkatú – bár vékony csontozattal – a chinoid ember is és ez az embertípus is már közel 100 évezrede Kelet-Ázsiában él. Ha a genetikai eredményekre alapozott időrend szerint hagyta el a modern ember Afrikát – azaz a jelen előtt (j. e.) 50 évezrede – akkor miként tudott az eurázsiai hidegebb éghajlathoz oly gyorsan alkalmazkodni, hogy a legidősebb modern emberi csontvázak már ezt a testalkatot képviselik? Meg kell jegyeznünk, hogy a modern ember előtt Eurázsia nagy részén neandervölgyi ember élt, akinek ugyancsak hideg övi volt a testalkata. A jégkorszakok során csak viszonylag rövid ideig volt a maihoz hasonló éghajlat, általában hidegebb volt és ezért a száraz, hideg éghajlati területek Eurázsiában mélyebbre nyúltak a mainál.
Az elképzelések szerint az ember a nagyemlősök családjából Afrika középső, keleti részén vált le, alakult ki, és járt be közel egymillió éves sajátos utat. Ez a terület az emberi élet során mindig a meleg éghajlati övezethez tartozott, azaz joggal várhatnánk el, hogy az ember alaptípusa a meleg övi. A már egyenesen járó, tüzet használó és szerszámait pattintó ősember, a Homo erectus, az archaikus ember viszont a günz jégkorszak idején már bizonyára elhagyta Afrikát és átkerült Eurázsiába. Ugyan a Kaukázus délkeleti területein a Homo erectus elődjének, a Homo ergatrasnak a nyomai is fellelhetők (másfél millió éves), mégis, az ember ‘tömeges’ elterjedése Eurázsiában csupán a günz idején történt meg. Szórványosan azonban korábbi emberi leletek is ismertek mind Európában, mind Ázsiában.26
A koponya legfőbb jellemzői: hosszúság, szélesség, magasság és a forma. A hosszúság a felülnézetbeli koponya legnagyobb méretét, a szélesség a koponya legnagyobb szélességét, a koponya jelzőszám pedig a szélesség és a hosszúság arányának a százszorosát jelenti. Alapvetően három típust különböztetnek meg: hosszú, rövid és közepes koponyát (koponyajelző kisebb, mint 75, nagyobb, mint 80, illetve 75-80).27 A koponya szélesség mellett az arc-, és a homlok szélességet is meg szokás adni. Ismeretesek széles és keskeny arcú típusok. Ez a koponya magasságra vonatkoztatott szélességet jelenti. Ezen belül is fontos a járomcsont, ill. az állkapocs szélessége. Fontos még a koponya alakja, azaz a homlok, a tarkó görbültsége. Vannak alacsony és magas, meredek, vagy lapos homlokú, vannak csapott, görbült, vagy csúcsos tarkójú embertípusok. Fontos még az orr formája, ami egyenes, konkáv és konvex lehet. A koponya magassága a fej magasságát jelenti. Alacsony és magas koponyájú típusokat különböztetnek meg.
Testmagasságot tekintve alacsony és magas típust tartanak nyilván. A test felépítése lehet robusztus és lehet vékony (filigrán, azaz latin eredetű angol szóval: gracilis). A testhosszúság arányában is vannak különbségek. Melegövi típusoknál általában a láb hosszú a törzshöz képest, a hideg övieknél ez fordított.
Nem győzzük hangsúlyozni, hogy élettani értelemben véve az emberiség egyetlen fajhoz tartozik, de ezen belül több alcsoportra (fajta, idegen szóval rassz) osztható fel. Általában öt alapvető csoportot különítenek el: veddo-ausztralid, europid (eurázsiai), mongolid, afrikoid és amerindid.28 Ez három eredetre is utalhat, jóllehet az eredet kérdése ma még nyitott.29 A magyarság szempontjából, az ötből kettőt kell figyelembe vennünk: az europidot és a mongolidot. Az utóbbi jellegzetesen hideg övi, azaz alacsony, rövid lábú, széles, lapos arcú típus. Ebben a vonatkozásban rokona a neandervölgyinek, aki szintén rövid lábú, széles arcú volt, de az arca nem volt lapos. Robusztus testfelépítésű, és vastag csontú a neandervölgyi ember. Kiszely meg is említi, hogy a mongolid is a neandervölgyiből eredhet,30 jóllehet a mongolid azért vékony csontozatú. Emlékeztetnünk kell arra, hogy a mongolid típus Kínában már a würm elején megjelent.31
A robusztus, vastag csontozatú ázsiai ősember a jávai ember, akinek fejformáját a későbbi ausztralid embernél vélik fölismerni.32 Velük szemben áll az afrikoid, ill. europid ember a hosszú lábbal, vékony, magas testfölépítéssel és hosszú, keskeny arcú fejjel. Az afrikoidot és az europidot gyakran összevontan kezelik, mert testfelépítésük hasonló, ámde az afrikoid orra lapos, széles, az europidé meg általában keskeny, kiálló. Az ausztralid típusú embernek is afrikoid típusú orra van, testfelépítése azonban más.
Az europid embertípusokat a mai ‘rendőrségi’ források általában kaukázusinak jelölik. Ez természetesen nem egyetlen embertípus megjelölésére szolgál. A klasszikus kaukázusi europid embertípus hosszú fejű, keskeny arcú, vékony testalkatú. Ezek közül a magas testalkat az indoeurópaiak és a nordikus emberek klasszikus jellemzője,33 az alacsony termet meg a keleti mediterrán emberé. Velük szemben állnak viszont a crô-magnonid típusok, amelyek általában hosszú és széles fejűek, széles alsó arcúak, robusztus testalkatúak és vastag csontúak. A crô-magnon A típushoz magas, a B típushoz alacsony testalkat tartozik. Az utóbbi rövidebb koponyájú változatát kelet-európainak (kelet-balti) nevezi Kiszely.34 Az európai földrész legrégebbi embertípusai ezek.35 Rövid, kerek koponyájú, meredek homlokú, kis közepes testmagasságú ember az alpi típus. Az alpi embertípus a würm jégkorszak végén (mezolitikum) jelent meg Európában.36 Az alpi típushoz közel áll a lappid. Igen rövid fejű, kifejezetten alacsony, igen széles arcú emberek.37
A keleti mediterrán típushoz vékony testalkat, hosszú, ill. igen hosszú fej és keskeny, lágyan kecses arc tartozik.38 Ennek három altípusa van, az iráni, a transz-káspi és az indid. Közép mediterrán típushoz tartozik a pontuszi, a gracilis mediterrán és az észak-afrikai. A káspi változat magas, a pontuszi kis-közepes, az iráni nagy-közepes39 testmagasságú. A nyugati mediterrán magasabb és kissé robusztusabb testalkatú, mint a keletiek.40 Vékony és alacsony testalkat, hosszú fej és keskeny arc jellemzi már az átmeneti kőkortól ismert protomediterrán típust.41
Az armenoid (taurid, elő-ázsiai42) szintén meglehetősen régi embertípus. Rövid, közepesen rövid, ill. kerek fejű, lapos homlokú, vékony és közepes termetű, a Kaukázustól délre általánosan elterjedt típus.43 Ennek és a mediterránnak a keveréke alkotja az akkád-sumér civilizáció embertípusát.44
Később alakultak ki keveredés révén az urálid, a pamíri, a turanid és a dinári (adriai) embertípusok. Valamennyit rövid, vagy közepesen rövid fej jellemzi. A turanid (Europo-mongolid45) kialakulását Lipták időszámításunk kezdetére teszi.46 A pamíri és a turanid középmagas fejű, a turanid széles, a pamíri közepesen széles arcú (járomcsontú), nagy-közepes, vagy nagytermetű. A turanid feje nagy, a pamírié kicsi.47 Az urálid típus szintén europo-mongolid,48 azaz széles járomcsontú, rövid fejű, de erősen hátrahajló és az arctól hátrafelé szélesedő koponyájú, alacsony arcú, mongolid jellegű49 – europid-mongolid keverék, mongolid túlsúllyal. A dinári típus közepesen széles, rövid, ill. túl rövid fejű, keskeny arcú, magas termetű fajta.50
Számunkra külön figyelmet érdemelnek a crô-magnonid típusok. Az A típus általában Nyugat Európában fordul elő, a B típus pedig Közép- és Kelet-Európában. Magyarországon a B típus főleg az Alföldtől északkeletre található meg, de általánosan elterjedt a Kárpátoktól északkeletre, közvetlen annak környezetében, valamint ettől északra. A szlávok és a finnek klasszikus embertípusa ez. Elképzelhető, hogy ez a két embertípus a kaukázusi és az őshonos nyugat-európai (neandervölgyi) ősember keveredésével jött létre.51 Jóllehet, egy ilyen keveredésre utaló konkrét archeológiai (csont) adatokat találhatunk,52 mások mégis hevesen ellenzik a gondolatot.53 Lipták a Kárpát-medencei embertípusok vizsgálatánál bevezette a crô-magnon C és C+ turanid típusokat is, de ezeket Henkey a turanidok közé sorolta.54
A crô-magnon B és mongolid típusok keverékeként jöhetett létre az uráli típus. A nordikus típus a klasszikus kaukázusi és a crô-magnon típusok keveredésével magyarázható.
Ki kell emelnünk, hogy a rövid és közepesen rövid fejforma jellegzetesen a Kaukázus környezetében, főleg attól Ázsia felé fordul elő, és föltehetően innen terjedt el más vidékek felé. A robusztus, hidegövi és emellett hosszú fejű embertípusok viszont jellegzetesen európaiak, annak is középső és nyugati területeire jellemzőek már az ősidők óta.
A honfoglalás kori magyarság embertani képét Bartucz és Lipták dolgozta fel. Kiszely elemzi a Kárpát-medence honfoglalás korát megelőző időszakai embertani képét is, ill. szembesíti a Kárpát-medencei embertani adatokat az eredeztetési modellek ún. őshaza helyszíneinek a megfelelő embertani anyagával.55 A elenkori magyarságról Henkeytől kapunk adatokat.56
A jelenkori magyarságot többségében a rövid és közepesen rövid fejű, széles arcú ember típus jellemzi. A testalkatot tekintve a közepesen magas alkat, inkább nehéz, mint vékony csontozat a jellemző. Ezzel szemben a környező u. n. indoeurópai embereket a vékony csontozat, hosszú fej és keskeny arc uralja. Egyik népesség sem homogén, egész Európa lakossága kifejezetten kevert típusú, csak az átlagok jelzik ezt a különbséget. Henkey a magyarság típusait Közép-Ázsia jelenkori lakosságával veti össze, akiket ugyancsak a rövid fej és a mérsékelten közepes testmagasság jellemez. Ebből ki nem mondottan arra következtet Henkey, hogy a magyarság eredete Közép-Ázsia. Azonban a robusztus csontozat, a széles arc és emiatt a közepes, ill. rövid fej nem föltétlenül kell, hogy elvigyen minket Közép-Ázsiába, ugyane a típusok lehetnek a Kárpát-medence jégkorszaki crô-magnon típusú embere utódai is.
______________
24. Lásd részletesebben Garn (1969) munkájában. Természetesen ma már a korábban még nem ismert immunológiai és gén adatokat is felhasználják, így a vércsoportot. Ennek megfelelően a három fő földrajzi alaptípust kilencre tágították fel. Ezek: amerikai, polinéziai, mikronéziai, malenéziai, ausztrál, ázsiai, indiai, európai és afrikai. Az általános kérdéseket illetően lásd még Kiszely (1976) munkáját.
25. Legújabb hírek szerint a 9 évezreddel ezelőtt a Cheddar egyik barlangjában elhalálozott ember génjeihez a legközelebbi génnel az ottani mai helység tanítója rendelkezett, ami arra utal, hogy bármiféle nyelvi változás ellenére a terület letelepedett lakói ott élhetnek már évezredek óta. Lásd: Science (1999).
26. Gáboriné (1980), p.: 80 említi a franciaországi Chilchac 0,5-1,8 millió éves telepét, Fagan (1989), p.: 125 a jávai 0,6-0,9 millió éves és p.: 126 a pekingi 0,4-2,6 millió éves emberről tesz említést.
41. Ez az ember élt a Shanidar barlang átmeneti kőkora idején Kr.e. 9000 körül. Valószínű emberáldozatot gyakorolt, Roux (1992), p.: 40. Érdemes ezzel szemben fölidézni az ugyanitt több tízezer évvel korábban élt neandervölgyi ősembert, aki nem áldozott embert, hanem a nyomorékot megmentette, megoperálta, életben tartotta.
Sz:
Magyarságtudományi tanulmányok, BP, 2008, 245. o
Magyarságtudományi tanulmányok, BP, 2008, 247. o
Magyarságtudományi tanulmányok, BP, 2008, 242. o